- 1월 7일 수업 하나만 놓칠듯 (1월 21일까지 총 7개)
- Zweikeimblättrige KulturPF :
- 시험 : 필기, 10개 + 3개 추가질문,
- 프락티쿰 : 실험한다, 온실 밀 관찰, 표현형관찰 Phänotypisierung - Weizenähren, Markeranalyse - 440개체 중 2개 마커 관찰, mRNA, in-situ Hybridisierung. 프로토콜은 간단히 쓸 듯.
[erste Vorlesung]
- 교짱 : Dr.Thorsten Schnurbusch von IPK. 최근 교수됨. 감자, 랍스 등 Getreide
- Praktikum : 10-14, 17-21, 24-28.02 셋 중 택 일 (12.11 까지). 프락티쿰 금욜 오전에 90min Klausur 있다. 최소 3명부터 가능하고
2. Geschichte : 히포크라테스, 아리스토텔레스,
- PFanatomie : Nehemia Grew, 식물세포 발견자 Entdecker
- Marcello Malpighi : 식물해부학, 비교생리학
- Zelltheorie : Matthias jacob Schleiden (세포이론), Theodor Schwann (Embryologie)
- Genetik : Mendel, Willhem L.Johannsen (Genotyp, Phänotyp)
5. Evolution der PF : Mono-, Dicots 로 일단 구분
6. Unterschiede : 디코튤은 뿌리가 크게 뻗고 주변에 나아가는 형태, 모노는 상대적으로 unstrukturell
- monocot : 잎이 그냥 파라렐. 씨는
- dicot : 스켈레톤이 있는 편. 씨앗은 카리옵져, 씨앗 차이는 나중에 다시 볼 것이다
7. Phylogenie der KulturPF :
8. Modellorganismen : E-coli, Hefe, A.t, Laubmoos (Algen ähnliches), C.elegans (Fadenwurm)
- höhere PF 조건 : Lebensperiode, kleines diploides Genom, bekannte Genom, zahlreicher Mutanten verfügbar,
- GefäßPF 조건 : kultivierbar..
9. Zweikeimblättrige KulturPF : 플로이드 짤을 볼 것
- 감자 : komplett Heterozykote, 즉 Heterosis 극대화. 하지만 클론이기에 Heterozygotgrad 를 유지하며 기를 수 있다
- Sojabohne : 문제는 이녀석 KurztagPF,
- Zuckerrübe : Kältereize 필요, Blüenbauen 은 in der Regel nie (?)
10. phylogenei wichtige : 쿨투어플렌쩬은 대부분 디코튤. 모노코튤 쿨투어는 상대적으로 moderner entwicklt (Gräser)
11. Phylogenie der Gräser : relativ neu. Gräser 가 인류에겐 최고다 이말이야
13. Gräser der Tropen : 하나하나 Genom bekannt
- Mais : 상대적으로 다른애에 비해 큰 Genomgröße,
- Mutanten verfügbar : klassischer Mutanten (페노튜피쉬 뮤탄트, 알려진 것. 표현형이 알려진 뮤턴트) 또 다른 건 뮤턴트인데 틸링같은걸로 의도적으로 만들었는데 누가 어떤 유전자형을 등록하면 그게 새로운 뮤턴트. 즉, 사람들이 아직 안써먹으면 뮤턴트 없다는거나 마찬가지
14. gemäßigter Zonen :
- Brachpodium distachyon : 아라비돕시스 같은 애란다,
- Weizen :
15. Speed Breeding : beschleunigen Wachstum, IPK에서도 해봤다고. 콘이 약간 얇고 쪼그라들었다고 schrümpftlich. 그럼에도 signifikante Bedeutung.
16. phylogenetische Herkünfte des heutigen Weizens : Triticeae
17. das Konzept der Kolinearität : Polyploidisierung 과 연관된 테마? Codon 중 단지 Exon만 해당된다!
- 앞의 Triticea 로 설명한다ㅏ.
- Verwandtschaftsgrad
- Homöologe : 밀의 A, B, D 각각은 호뭬올로기 관계
- Orthologe : 같은 Vorfahren
- Paraloge : Genduplikation 의 결과 예를 들면. A - A´
https://terms.naver.com/entry.nhn?docId=125349&cid=43658&categoryId=43658
18. Genom-Kolinearität : 엑손을 의미한다구
19. Genom-Kolinearität zwischen Weizen und Reis : 붉은 부분은 높은 콜리네아리텟을 보여준다는 걸 알 수 있다
23. Das Pan-Genom : 의미는 일단 mehr als ein Genom
- Summe aller Gene aller Individuen
- Kern-Genom : 503 개 인쭈흐트리니에 옥수수 중 완전 겹치는 건 오직 20프로.
- entbehrliche Gene : 이미 사리진 유전자도 포함하는 듯. 위 사례에서 80프로. 생각보다 너무 이게 많았다 이말이야
- synthetische Biologie와 관련있는 분야. 어느정도의 유전자가 그 생물을 생물로서 유지하기 위한 최소한인가?
24. das Gen-Konzept :
25. Mechanismen der Genduplikation : 온갖 가능성들 젱장
26. Entdeckung der jumping genes : 바바라 맥클린톡
27. jumping genes : 정크 진의 Variation이 엄청 많은 이유는 역시 식물은 뮤턴트 가능성을 많이 가져야 살아남을 수 있기 때문인듯
28. Mutationen lassen Gene evolvieren : Sequenz가 어떻게 변화하는가
29. Mechanismen der Genevolition : 이기적 유전자
- 엄청많다...
- F. Neofunktionalisierung :
30. Fluß biologischer Information :
31. Vom Gen zum Protein : 여기서부턴 상대적으로 기초
33. Regulation eukaryotischer Genexpression : Transkriptionsfaktor,
34. Transkriptionsfaktoren : 아무 중요하다 이말이야
- unterschiedliche DNA-Bindungsdomänen : 크게 3개 있다고.
- leucine 은 알파 아미노산의 한 종류
36. Erfindung der Landwirtschaft : menschlich말고도 Ameisen 이런거.
- aktiv Kultivierung zur Begrasung,
- 아마, 밀, 쌀처럼 흔히 자연에 널린 애들을 기르기 위한게 아니었다고 생각. 오히려 자연에 wenig vorkommen 하는 종을 기르기 위해 인공적으로 시작했다고 생각. z.B) Linsen. 집 앞에서 좀 기르고 하면 훨씬 나은 Nährwert 가질 수 있을 거라고.즉, Getreide-PF 이 시작이 아니라 다른 종류의 식물이 아닐까 한다고.
39. Domestikation von KulturPF : lange komplizierte Prozess
41. Domestikation verläuft stufenweise : Domestizierung의 시작 (좀 멀쩡하고 먹기 쉬운 애들로 골라 길러진다) -> 새로운 다양한 종자 -> kultivierter Formen 다양한 환경, 지역에 적합한 애들 -> 방향성을 가진 PF-Züchtung
42. Domestikationsmerkmale von KulturPF :
43. fühe Domestikationsmerkmale : 콘 크기, 형태, Spindelbrüchigkeit - 이게 아마 가장 중요한 요소일 듯. 우리는 더 강하게 붙어있는게ㅔ 필요하다. Wildgerste는 너무 쉽게 leicht fällt
- Samen und Frucht 차이 : Endosperm und Ka? 같이 있으면 Frucht라네??
44. erste Gene für Spindelbrüchigkeit in Geste : Udda Lundqvist 이 분이 40-60년대 수많은 보리 뮤턴트 만드셨고 아직도 쓰인다고. 일명 보리 레전드.
- btr 무타찌온 하나만 있어도 Spindelbrüchtigkeit 가 약해진다 -> 더 강하게 붙어있는다
47. erste Gene für Spindelbrüchigkeit (brittle rachis) : btr1 oder btr2 Mutation 은 대략 만년전 발생한 것 같다고.
48. parallele Domestikation / Selektion bei Korn- / Samenabwurf : Sorghum, Mais, Reis에서의 낟알이 떨어지는 메커니즘. Spindelbrüchigkeit는 상대적으로 특이한 경우다
49. Grundsätzlicher Aufbau eines Ährchens bei Gräsern :
50. Domestikation von Mais : TGA1 von Lysin(K) zu Asparagin (N) 이게 핵심적 역할
51. Domestikation von freidreschender Gerste : hulled and - Naked Caryosis 껍질이 훌렁훌렁
52. Dinkel은 좀 다른 케이스
53. Bespelzter Dinkel : einzige Ausnahme
- q Gen : Q 만 있어도 dominante.
54. Dinkel- eine Kreuzung aus Brotweizen und Emmer
55. Auswuchfestigkeit (Dormanz) bei Getreide : 이것도 아주 중요하다 이말이야
- Auswuchs 라 하면 Samenruhe 가 깨지는 것을 말한다. 날씨 등의 요인으로 Keimung이 가능한 형태가 되는 것.
- Samenruhe 를 말한다. eine WildPFmerkmal
- https://terms.naver.com/entry.nhn?docId=5646750&cid=62861&categoryId=62861
56. erste Allele für Auswuchfestigkeit (Dormanz) - Dormanz zu vermeiden
- Qsd1 kodiert für eine Alanin-Aminotransferase - 2016 일본에서 연구
57. Weizen-Allele des Qsd1-Gens - 역시 위와 같은 일본 연구팀
58. andere Gene für Dormanz
59. 마지막 : 어떤 애들이 중요했나요 뭐 이런거.
* Primordium development in plantsPlants produce both leaf and flower primordia cells at the shoot apical meristem (SAM). Primordium development in plants is critical to the proper positioning and development of plant organs and cells.
- infloreszenz
-
[zweite]
- Domestikationsmerkmal 을 다룬 이유 : 아마 이 사례들이 Entwicklungsgenetik의 목적에 제일 부합하는 모범사례
- 복습 : btr1,btr2, sh1, tga1, nud, Q, tg 각 어떤 Merkmale 에 해당하는 Gene인가 - Spindelbrüchigkeit등
- 복습 : 우리와 KulturPF 의 관계는 상호의존적이다. 우리는 당연한거고 식물은 이미 Merkmal 이 Kultur에 고정된 경우 Spindelbruch를 위해 인간의 도움이 필요하다. 즉, 인간이 없음 이녀석들 Fortpflanzung 안될걸
1. Zweizeiligkeit bei Gerste : Zeiligkeit 의 으미는 Anzahl der Körnerreihen pro Ähre
- 보리는 양쪽이 동일하게 나는 것 같아도 사실은 번갈아 나는 형태.
- Zweizeiligkeit : pro Spindelstufe 3개의 Blütchen이 달려있으나 중간것만 fertil하고 둘은 steril. 이건 전형적은 Braugerste. niedrig Protein 이지만 인간 영양을 위해. 그리고 아무래도 하나만 있는 편이 동일한 퀄리티로 가공하기 쉽다.
- Sechzeiligkeit : vrs1 뮤턴트를 이용해 저 Steril 꽃 2개도 fertil하게 만듦 -> 3다 알이 생긴다. 이건 프로틴이 많아서 Schweinfutter로 쓰인다.
- Vrs1 als mRNA: Primordien 은 식물의 기관(Blatt, Blüte, Wurzel) 이 형성되는 조직을 말함. 이 녀석을 중심으로 Embryo-Ähren을 보면 Lateralblütchen (옆에 붙은 작은 꽃) 의 Fertilität 과 연관.
- 디테일 : Lateralblütchen 이 verzögert -> 꽃 내에 Fruchtknoten 이 형성되지 못함 / Anthere는 3다 형성되어있다 비록 발달이 늦었더라도!
2. Evolution von Vrs1 : Hox2, Hox1
- Hox2, Vrs1 단백질의 구조는 아주 비슷하지만 Expression 위치는 꽤 다르다.
- Proteininteraktion : Zip 은 프로틴과 프로틴을 붙여주는 아인가보다. 각각의 프로틴은 TF에 붙어서 억제? 아래 B 설이 더 현실적이라는데 둘이 같은 Tf 를 두고 경쟁한다는 것. 기본적으로 둘은 매우 비슷하므로.
3. Vrs1.t (Deficiens) Allel : Vrs1 에서 몇몇 Allel이 바뀌면 이렇게 양쪽 Blütchen형성은 압도적으로 억제 (전자현미경에서 확인가능) 되고 Zentral Korn의 Protein이 더 커진다. umgenutzt werden이라고 말하네
- 두개 이름이 다른 애는 아마 앞이나 뒤에 AS다른 부분이 있을거라고.
- UK, Schortland 에서 Def2 는 zugelassen 되어서 굉장히 성공적이었다고
- 이 Kreis의 크기는 클수록 오른쪽 위의 종들과 Haplotype 의 공통점이 크다고. 중간의 15.2는 엔세스트랄. 거기서 Vrs1.b3 SNP 가 이제 처음 쓰임새있게 뻗어져나왔다고. 또 여기서 봐야할 것은 엔쎄스터로부터 unabhängig한 상태로 Sechszeilige 가 생겨났다는 것이다. 그런 진화적 배경은 우린 오직 spekulierbar = nicht beweisbar. 한 설은 야생종으로 부터 6줄 생겨났는데 이게 더 무거워서 땅에 더 가까이 많이 낟알을 떨어뜨렸다고
https://terms.naver.com/entry.nhn?docId=5141387&cid=60266&categoryId=60266
4. GNI-A1 Lokus : 2019년 punbliziert. Weizen VRS1 ㅇㅣ다. VRS1 의 Ontologe grain number increase 1
- hohe Blütschenfertilität bei Weizen : Durum에 있어서 이 뮤턴트 (T 로 나타나있는 SNP) 가 셀렉되어 유독 두룸이 낟알 수가 많다고.
- GNI1 -RNAi 를 통해 Hexaploid 의 A, B, D Genom 모두의 GNI1 을 억제. Rachilla 가 길어지면서 다른 낟알도 달릴 수 있었다 이말이야
- GNI1 Allel in 210 hexaploid : 이 Allel이 Züchtung에 알려지고 적용된 이후 추이. 필드에서 10-30프로 Ertrag 향상을 가져왔다!! 즉, Kornzahl pro Ähre 는 sehr wichtige Ertragskomponente. 근데, TKG 에 있어서는 차이가 없다! (에어트락에는 당연히 유리하다 이말이야)
- 메커니즘 : VRS1 과 같이 negative Regulator für Lateralentwicklung. localisiert in apikalen Blütchen + Rachilla. Ähre끝 부분에서 주로 작동해서 꽃 absterben 하는 대신 주변의 아래 Korn은 발달시키게.
5. Blattphänotypen Vrs1 : 잎에 있어서 Vrs1 유무에 따라 차이가 난다구!
- Vrs1 뮤턴트 (6zeilige) 는 잎도 더 넓다. Blattader (vein, 잎맥?) 도 훨씬 능ㄹ어났음을 볼 수 있다
- 결론 : Vrs1 은 Ähre 뿐만 아니라 식물의 Blattmeristem 에도 negative Regulator 로써 작용함을 알 수 있다.
6. Übergangsformen zwischen zwei- und sechszeiliger Gerste : 확정적인 뮤턴트 외 quantitative Genetik과 연관있을 법한 여러 Variation이 natürlich vorhanden
- int-c Allele : wichtig für Fertilität und Ertrag
- int-c.b Allel 은 Fertilität 줄인다, c.a 는 Fertilität 올리고.
7. Lebensphasen : 2가지 - vegetativ (Spross Apikal Meristem) + reproduktiv
- A.t : 2개의 reproduktive Meristem 갖는다. 메리스템은 크게 1) apikale Primärmeristeme (SAM, WAM), 2) nichtapikale Sekundärmeristeme (Cambium, ...)
- Meristem : Stammzell이란 말이지. 여기 나온 식물 구조 볼만함. Primordien 은 앞으로 특정 기관으로 형성될 meristem 위의 특정 bereich 를 의미.
8. 어떻게 IM (infloreszenz-Meristems) 가 Blütenbildung으로 전환될까?
- 크게 2가지 Faktoren / Signale 로 나뉜다.
1) Exogenen : Tageslänge (Photoperiod), andauernder Kältereiz, T, Licht
2) Endogenen : GA, Zuckerstoffe, autonome Blütinduktion
* Tageslänge 설명할때 왜 유럽선 쌀이나 콩이 까다로운지 다시 언급함 (2번 언급됨). 기본적으론 Langtags 녀석은 여기선 힘드니까 neutral 한 sorten이 필요하다 인듯. 근데 밀도 장일인데?
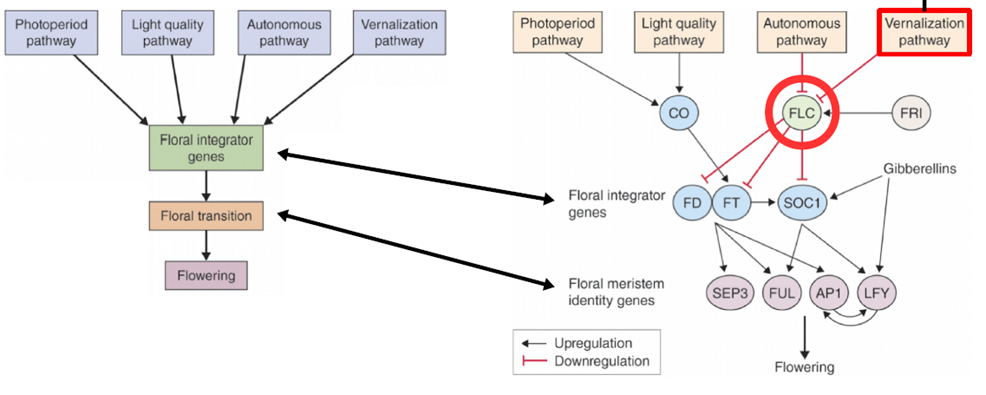
- Flowering Locus T (FT) : Ausgangspunkt.
* 예전에 다뤘던 것과 같은 Photoperiode Mechanisemen 인데 너무 각각의 단계마다 실험적 증거를 보여주느라 한번에 내용을 인식하기 어렵다. nicht gut organisiert. 좀 더 개괄적인 내용을 먼저 소개할 필요가 있는듯. 다만 최신 논문 (3년 내) 자료를 사용한다는 것과 Domestikationsmerkmal 과 같은 성공적인 Muster를 중심으로 발달에 주요한 Gene 를 다룬다는 건 수긍하게 되는 구조.
- miRNA 172 : FT 조지는데 주요한 역할
- Meristemsonen mit Genexpressionen im teilungsaktiven IM :
9. Umweltbedingte Faktoren der Blühregulation in Getreide : VRN,
- Apex (끄트머리) 에서 SAM -> IM 전환을 확인할 수 있다
10. Regulation der Blühinduktion in Getreide (gem. Breiten) Vernalisationsgen - VRN1 이 양반들이 처음 분리.
- Eam 종류의 역할 : Blühinduktion unabhängig von Ppd (Photoperiod). 즉, 이녀석들로 인해 낮과 밤에 일정하게 Ppd 영향을 주기적으로 준다. 하지만 뮤턴트로 꺼지면 매번 일정하게 준다.
- Blühzeitregulation in Gerste über Tageslänge : ppd 뮤턴트(ppd-H1 insensitive Allel) 는 보다시피 꽃피는 타이밍이 달라진다. 우야등등 이런 변화를 적용하려면 어데서 키울지가 중요하다. 왜냐면, 아무리 해가 길고 따끈한 곳이라도 너무 빨리 RM 으로 전환되면 Biomasse 가 적은 상태에서 만들어야되기 때문.
- 이런 연유로, 모든 Sommergerste in mittel Europa 는 ppd-H1 Allel 을 갖고있다.
- 아래 표에서 왼쪽 푸른색과 오른쪽 연두색의 Gene를 적절히 조합해야 각 지역에 맞는 식물을 골라서 기를 수 있다. 그 예시로 이 Anpassung이 없이는 스칸디나비아에서 보리 키우는건 무리였을 것이다.
- Hexaploide Weizen은 Ppd-D1, B1, A1 각각 페어마다 하나씩 Ppd gene 갖고있다.
- Norman Borlaug 의 Dwarf wheat 는 단순히 키만 작은게 아니라 neutral Photoperiode 도 갖고 있었다.
- 다음 수업 : 어떤 Meristem 의 gene 이 Entwicklung에 주요한 역할을 하는지, 식물 발달에 관해 얘기할 거임.
-
[그다음 수업]
1. Was ist Entwicklung : KulturPF 은 좀 복잡한 관계로 모델 플란쩬으로 한번 훓어보고자 한다.
- Tabak, At
- 중요한 Phasen : Gametophytenentwicklung, Embryogenese (둘다 nicht sichtbar), Keimung, Blütenbildung
- Entwicklung 은 기본적으로 Morphogenese 형태 변화
1) Differenzierung(특정 기관 분화), 2) Morphogenese(형태 이룸), 3) Wachstum (양적 증가)
- Entwicklung의 Grundlage 는 Zellwachstum (1. Zellvermehrung oder Teilung, 2. Zellvergrößerung - 이건 Regulation의 관점에서 식물의 세포벽과 연관해 wichtige Aspekt)
2. Entwicklung - Differenzierung : das Verschiedenenwerden
- Ursachen : 1. chemisch, elektirisch, mechanisch (호르몬 등), 2. Gene und Genexpression, 3. zur Musterbildung - 그 스스로도 구조적 원인이라 치는 듯 (engl. patterning), 4. über Zell-Zell Kontakte (Erkennung der Bakterien), 5. Positionsinformation und - interpretation (시그널이 잎에서 뿌리로 가고 이런거)
- Definition - Morphogen : Signalmoleküle (Hormon, Protein)을 의미. 기본 원칙은, 농도차에 의해 모든 분화가능성을 가진 세포 내 Schwelle 도달에 따라 다른 구조로 변화
* on, off 수준의 Schwelle 를 가진 메커니즘은 없을까?
- 세포분화는 Resultat differenzieller Genexpression
- 모든 Zelldifferenzierung bzw. Determination 은 PFzellen에선 reversibel (Totipotenz) : 그걸 이용한게 in-vitro kultur. 이 의의는 벌써 분화한 세포도 Zygote로 돌릴 수 있다는 것 (진짜??)
* 잘은 모르는데 이걸 이용하면 화공처럼 우수한 품종을 대량으로 클론처럼 만들 수 있다는 거 아님? 꼭 감자아니라도?
- Zellteilung 구분 : Perikline, Antikline (위아래, 좌우)
- 동물과 식물 차이 : 식물은 postorganese 즉, 다 크고도 추가적 기관형성이 가능. 사람은 안대여 간을 하나 더 만들 순 없다. (자연적으론, 그리고 아직은)
- Sproß- und Wurzelapikalmeristem : SAM. WAM. 뿌리, Sproß 경계 구분은 Auxin 농도 구배에 따라 결정된다 (구분이 시작된다).
3. SAM : 여기서 뭐가 일어날까? SCN을 중심으로 분화되면서 양측으로 분화된 세포양이 늘어난다. 그러면 중앙의 SCN은 위로 솟아오를 것. WUS, CLV3, HEC1 은 이 과정을 보여주는 대표적 예시 (각자 프로틴). 그리고 Wus-CLV3 는 영향을 주고 네거티브하게 다시 피드백이 간다. 그당므 그림의 HD-ZIP 는 TF. 하나 중요한 건 Sproß에서는 Auxin이 Zelldifferenzierung induziert,
- SAM Hormon 메커니즘 : 옥신과 씨토키닌 의 역할 및 분포를 잘 보자꾸.
- CLV-WUS 메커니즘은 아라비돕시스에서 잘 연구되었고 모델이 있는데, 이게 Reis, Mais 에서는 전혀 다르다. KulturPF에서 호몰로그를 관찰해봤는데 달랐다고. 하지만 염두에 둬야될 게 식물이 Apikale Dominanz를 이뤄가는 방법이 아라비돕시스 형태만 있는게 아니란 것을 신경쓰자
- Mais 에서 보는 SAM 책임자 유전자 : FEA3 뮤턴트는 보다시피 위로 가기보다 옆으로 퍼진다. B73, W22는 inzuchtlinie고 그걸 하이브리드 해서 Heterosis 이끌어낸 fea3 는 훨 많은 körner
- Fasciation (Fasziation) : 기관이 breite 하게 커지는 Mutant. Ursache - kormonelle Umsteuerung meristematischer Zellen, Mutationen, Befall oder Umwelteinflüsse (Wuchsregulatoren) 질문 Gallen 도 그럼 이거?
- Cell Fate Map : 싱기방기
- Cell fate map in At : positionsabhängig SAM 세포의 운명은 시작 시점에서 어느 위치에 있냐에 결정된다. 여기 나타난 숫자 그대로 분화된다. 그렇기 때문에 위치 결정적이라 말하 룻 있다.
- fate map über Zelllinien und Chimäre : 여기 나타난 점 뮤턴트는 그 뮤턴트가 후에 딸세포로 전달되고 하면서 궁극적으로 이렇게 그 영역은 전부 뮤턴트의 영향이 발생한다. 아마 이런 발견은 자연적으로 봤던 걸 방사능 노출 같은걸로 실험해서 밝혀낸 다음 아하! 했을 거라는 듯.
- Stammzellenerhaltung, Zellteilung, Differenzierung und Organanlage :
- Homeobox 유전자 :
- Eudicot und zweikeimblättrige 의 차이
- C3, C4 식물 차이 및 기후변화에 관해 장단
4. Organanlage, Boundaryformation und Phyllotaxis (잎차례, 잎이 달리는 순서)
- Organbegrenzungsbereich : 바운더리. 이 바운더리의 존재로 인해 (Grundvoraussetzung) 식물은 nicht zur Verwechselung kommen , 성장할 기관의 위치를 자리잡게 된다. 그림에 보이듯, 플라스모디움이 이 그렌쩨 사이에 발생하여 추후 잎 이란 기관으로 분화, 생장
- Homeobox TF Mais KNPTTED-1 : spezifiziert Sprossmeristeme 이거 2000년 초반 발견. 제일 왼쪽은 mRNA expression 위치, 오른쪽 그림은 프로틴.
- mRNA in situ Hybridisierungssignal : 바운더리 유전자 AtLAS (lateral Suppresor) 관찰.
* 흥미롭다! 이 구분을 통해 뭐가 생겨날지 결정되는 것 처럼 보임.
- Formation eines Grenzbereichs : 바운더리의 중요성이다 이말이야. 바운더리는 굉장히 langsame Wachstum 으로 gekennzeichnet. 이 구간의 Merkmal이라 할 수 있다
- Hormon- und Genregulation von 바운더리 formation : CUC외 다른 유전자, 호르몬의 농도구배가 ab 한다는 걸 알 수 있다.
- wichtige Gene für Boundary : 여기 이것저것
- Phyllotaxis : Grundanordnungen der Blätter oder Blütenstände am Spross :
* goldene Schnitt 황금비 : P0, P1 플라스모디움 간의 Winkel 은 항상 137.5도 (베르누이) 를 이룬다! 단순히 아라비돕시스 뿐만 아니라 자연꼐 온갖 것에서 다 발견. 원인은 아직 모른다?
5. Blattentwicklung : 우린 과학자니까 구체적으로 잎을 알아야 된다고...
- 옥신이 Entwicklung에 관여 안하는 곳을 찾기가 힘들다고.
- Etablierung der Blattspreitenpolarität : 잎 oberseite / unterseite 의 구분을 자아내는 것. 잎이 제대로 성장하기 전, 아주 이른 스테이지에서 부터 Adaxial, Abaxial 로 위 아래 의 구분이 생긴다. 그 다음엔 여기 보이듯 온갖 요소가 위 아래 다르게 영향을 미친다
- Erhalt der Blattspreitenpolarität durch Erkennung eines Schwellenwertes für mobile miRNA Gradienten : 2017 논문에서 WT 과 달리 tasiARF 를 Umsteuerung 하니까 잎의 ober, unterseite 구분인 Polarität이 깨졌음을 발견.
- Vaskularisierung (Gefäßbildung) : 잎 내의 베스큘러 시스템을 갖추는 데 Auxin 이 주요 역할. organ은 스스로 Gefäßsystem 을 bilden 해야하지만 동시에 Stamm과 연결될 Hauptgefäß와도 연결이 되어야 한다. 옥신이 이런 과정에 주요 관여한다.
- Prokambium : 향후 Phloem, Xylem 으로 발달할 원초 베스큘러 슈탐쩰.
- Adulte Blattformen 쌍떡, 외떡잎식물 비교 : c Mais는 다른 쌍떡잎과 달리 au 구조 가진다.
- Grasblätter : 대로 보이는 Blattscheide, 와 멀쩡한 잎으로 보이는 Blattspreite 둘 다 합쳐서 Blattorgan. Blattscheide를 보면 안쪽이 jung, 바깥이 올드한 잎대. 그리고 노란부분은 세포가 많아지는거고 초록색 부분은 세포 크기가 늘어나는 것
- zweikeimblättrige 에서 다양한 잎 분화 모델 : CUC2는 바운더리 유전자이고 이게 분화되는 곳 = Zellverteilung 아주 낮다. 이런 바운더리 포메이션의 특정적 -> 다양한 잎 형태
- Anastomose : Querverbindungen zwischen Blattadern (leaf vein, 잎맥)
- Spaltöffnungen (Stomata) : Gasaustausch (CO2, O2) 가 일어나는 곳, Stomata 닫힘은 K+ 이온이 증가해서 물이 들어오면서 이뤄진다.
- 스토마타 형성 : 시작은 protodermal cell. Meristemoid 단계에서 symetric 한 구분이 이뤄지는데 이게 핵심 보인트. 꽤 잘 밝혀진 klarer linialer Weg
- Stomata bei Gräsern : 앞의 많은 예시들과 달리 4개의 Schließzellen로 이뤄진다. 관여하는 TF 는 아라비돕시스와 유사.
- Photosynthetischaktives Organ : C3, C4 차이 깔끔하게 정리한 그림. C4 는 C3 로부터 발생했다! C4는 hell trocken에 많이 발견되고, C3 는 schattig feucht 한 열대 에서 많이 발견. C4 는 hohe CO2 Aufnahme 를 위해 hohe T 필요하다.
- Entwicklung der C4-Kranzanatomie bei Mais : C4 에서 발견되는 구조.
- C4 Rice : 빌 앤 멜린다 게이츠 재단서 후원 하에 진행중. 독일은 뒤셀도르프에서 3, 4개 그룹이 참여중이라는 듯.
* 책 추천 :plant transcription factors - gonzalez 2016, 박사하려면 아마 필요할 거라네.
- Achselmeristem :
- Premodium 은 잎의 전구체인듯
- Überexpression : 정확히 다른 expression과 구분
- Halmen :
[Phytomer- Konzept]
1. Sproßaufbau über Phytomere : Phytomer는 Grundmodul für Sproßentwicklung. Intermodium, Knoten, Blatt, Achselmeristem 으로 이루어진 성장 파트를 말한다.
- Infloreszenzmeristem : 초록 링은 Blattmeristem, 노란색은 Achselmeristem이다. 이 아으젤메리스템이 후에 Ähre로 성장할 가능성을 가지고 이 단계에서 Blattmeristem은 이 단계에서 성장이 unterdrückt.
- in Gräser : 중앙하단의 단위가 기본적은 Phytomer. 아래는 Bud (Knospe) 와 뿌리 성장 가능성이 있고, 위로는 잎. 오른쪽 그림을 보면 Knoten (node) 를 기분으로 퓨토머 1, 2 의 구분이가능핟. 즉, 한 크노튼은 2개의 퓨토머 단위가 합쳐서 이뤄진다. 왼쪽 상단을 보면, vegetative / reproduktiv 타입의 퓨토머 차이가 그려져있다ㅏ. Collar 는 Ähre가 달릴 수 있는 basis 라고 생각하면 됨 콜라는 써프레쓰드 플랏플랏츠. 한 Ährchenmeristem 이 한 Ähre 위치에 존재. 한개씩.
- in Gräser : Phytomer 컨셉에 따라 각 기관이 퓨토머로 부터 어떻게 entstanden하는지 그려져있다. Lemma 는 잎이 달리는 기본 위치. 오른쪽 Weizen을 보면 왼쪽 보리와 다르게 여러 꽃이 한 Spindelstufe에서 발생
- im Gräser : Seminalwurzel - 뿌리가 자란다 , Nodalwurzel - 여기선 Knoten 이 자란다. 핵심 그레저에서 뻗어나온 Bestockungstrieb은 Achselmeristem에서 유래된 것.
* Samen 안에서 사실 이미 4-5 Blätter의 가능성을 가진 Plasmodium이 있다. 그 말은 이미 거기서부터 Phytomer의 존재를 생각할 수 있다는 것. 그리고 여기서 Seminalwurzel이 생기겠지. 하지만 그 위에서 생기는 Phytomer는 전부 Nodalwurzel 을 생성.
2. Hormonelle Steuerung während der Sprossentwicklung : Auxin (IAA), Cytokinine (CK). Strigolaktone (SL). 옥신은 Pin Protein (옥신 트랜스포터) 를 통해 Phloem으로 이동, CK und SL은 Xylem 으로 이동. 옥신은 Apikal-Meristem에서 생성되고 CK, SL츤 Wurzel에서 생성된다.
- SL : Antiverzweigungshormon
- Regulation der Großentwicklung : MAX1,3,4 는 SL Biosynthesegene 이고 MAX2 는 시그널 - Root가 WT라도 shoot 가 max2 뮤턴트면 안됨!. 실험의 오른쪽 끝을 보면 뿌리는 WT, Sproß는 mutant인걸 붙여서 길러보니 WT처럼 멀쩡하게 자랐다고!!
- Apikale Dominanz ist durch lokale Zuckergehalte im AxM reguliert : 당이 많이 쌓인것 자체가 Apikale Dominanz brechen 을 위한 시그널이다.
- Lateral surpressor in At und ;onoculm 1 in Reis : 서로 orthologe
- 옥수수의 Domestikationsmerkmal 중 하나 : Reduktion der Seitentriebe - tb1 Gene (TF aus der TCP-Familie). 그다음 페이지 TE - 62kb어쩌고 보면 cis-regulatorische Region. 어떻게 오른쪽 그림에서 이 구간을 찾았지?
- tin1 을 보면 Standfestigkeit와 연관있다고 : UTR 과 관련된 녀석이다. 결론적이로 이게 다른 Gene의 Expression에 영향을 준다. UTR은 자주나오네.
- Auxin - CK - TB1 - GT1 의 메커니즘이 최근의 모델. CK는 Wurzel에서 생성된 다음 TB1에 작용하지만 그다음에 이게 뭐 분해될지 어떨지는 모른다고. 이 작용도 아마 indirekt일거라고.
3. Internodverlängerung - und verkürzung
- Schossen : Verlängerung der Halminternodien. 마지막 Internodium은 Ähre전의 노드이며 peduncle 라 불린다. 각 Phytomer의 윗부분이 늘어나는 Meristem - 파란색으로 표시됨
- Internodienentwicklung 의 Musterbildung은 아직 정확히 아주 정확히 밝혀지진 않았다! 관련 질문은wie , wann bilden sich die Zonen, 어떤 유전자가 Zonenidentität, 어떻게 Wachstumgsphase 에서 reguliert? 그리고 모든 Internodien이 gleich한가!
- Norman Borlaug : 작은 일본산 밀과 큼직 노말 밀을 제대로 섞기위해, 거진 15년간 Züchtung을 진행했다. 거진 30 Zyklen 을 반복한 것.
- Halmlänge beim Weizen : REDUCED HEIGHT (RHT1)-Gen. 이 유전자는 GA과 연관있으며 DELLA-Proteine 가 GA 의 Repressoren로 작동. 델라 Mutant는 dominant 한데,
- SEMI-DWARF1 (SD1) beim Reis : 아까거랑 하는 짓거리가 좀 비슷허네.
- BRASSINOSTEROID-INSENSITIVE1 (BRI1) der Gerste : 밀, 또는 쌀의 orthologie von RHT1, Della 를 뮤턴트해도 똑같이 positiv결과 안나왔다.
4. Blütenentwicklung
- At hat zwei reproduktive Meristem : IM und BM. 여기 그림 At 꽃 구조 설명으로 아주 (독어, 영어) 좋다!
- FM-Initiation über Auxin in Spiralanordnung : 예전에 말했던 Goldene Schritt. 이 그림 본 적있다.
- Blütenentwicklung bei Arabidopsis unterscheidet zwei Hauptphasen : floral primordium 이 생기고 그다음 floral organ이 발생하고.
- At 의 구간 별 FM, IM 구분 및 deren codierte Genen : 아라비돕시스 답게 무지하게 많이 연구됨.
- Fruchtknoten은 가장 최후에 differenziert되는 Stammzell 구간.
- ABC Modell : homöotische Gene (Homöosis : Transformation der Organidentität) - Expression am in verschiedenen Kombination führt zu Veränderungen in der normalen Entwicklung.
Umwandlung einer Körperstruktur in eine andere (homonome) Struktur, die sonst im Tier weiter anterior (anteriore Transformation, ö vgl. Abb. ) oder weiter posterior liegt (posteriore Transformation)
출처 : https://www.spektrum.de/lexikon/biologie/homoeose/32461
- ABC Gene는 사실 E Gene 인 Sepalate Gene의 도움이 필요하다!! 나중에 알려진 듯.
- ABCDE Modell : D Klasse gene 는 nur für Ovulenentwicklung
5. Makroentwicklungsstadien bei Getreide - BBCH (Zadoks?)
- Makrostadien 은 BBCH (Zadoks) Skala 로 식물다나 나누어진 구분 기준이 있다. 말그대로 마크로 슈타디엔.
- 그림으로 나타낸 예시가 한무더기. 이런 형태를 보이면 얼추 그 슈타디엔에 있다고 생각할 수 있는거임.
6. Blütenstandarchitektur : KulturPF마다 가지각색 꽃대 형태가 있다. 옥수수, 벼, sorghum, 등등
- Evolution의 과정에 따라 Ur-Form이 Ähre (잎대에 딱 붙어서 Spike달리는것), Rispe (약간 체리처럼?) 이런 형태로 분화됐다고 생각.
- Konzept der wechselnden Meristemidentitäten : 이거 언제 한번 나올 줄 알았다. At는 꽤 단순한 형태, 다른 Hauptgetreide는 아따 난리도 아님.
[그다음시간]
4. Konzept der wechselnden Meristemidentitäten : Gräser는 At에 비해 Meristem이 좀 더 복잡다.
- VM : vertical shoot meristem, IM : Inflorence (reproductive), FM : Floral
- BM : Branch Meristem, Verzweigung하는거
- SM : 이건 Gräser에만 있는 특수한 경우
- Weizen : Sonderheit가 있는데 FM 가 한 Ähre안에 여러개 생기는것. 이게 다른 애들에 비해 높은 Ertrag의 비결
5. Blütenstand
6. Weizenerträge & Kornzahl in DT : 밀은 높은 dicht 를 갖고 길러지는데 이런 환경에서 더 높은 Ertrag을 위해 더 꽃을 많이 피우는건 결코 쉽지않다.
- TKG 와 Kornzahl은 보다시피 negative Assoziation! : 각자와 Ertrag의 관계를 보면 TKG는 Ertrag에 Kornzahl에 비해 큰 영향을 주지 않는다. 과거 100년 전의 밀과 지금의 밀 TKG를 비교하면 큰 차이가 없다! 그래서 Züchter는 중간정도 크기의 콘을 유지하면서 더 많은 콘짤을 추구하게 된다. 아주 중요한 포인트
7. drei reproduktive Meristem : IM, SM, FM
- 각각의 Ähre는 서로 다른 Entwicklungsstadien에 놓여있다.
8. Lebenszyklus :
- collar initiation 에서 Terminal spikelet 에 도달하면 여기서 Anzahl der Ährechen이 결정된다.
9. Weizen . Infloreszenenentwicklung : 터미널 스파이클렛에 도달하면 더이상 스파이크 Meristem생성이 안된다
10. vegetativer Apex : 잎이 생기는 부위가 오른쪽 그림서 보인다.
12. double ridge (DR) : IM은 항상 apical 에 위치한다.
- Blütenbildung은 사실상 modifizierte Blätterbildung 과 다름없다.
14. Lemma primordium (LP) :
15. Terminal spikelet (TS) : 꽤 중요해보이는구마
- 그다음 검은 사진에서 숫자를 볼 수 있다. 제일 위가 융하고 아래가 오래되서 Entwicklungsgradient라고 칭할 수 있다.
- 매번 스타디엔을 설명하는 그림은 앞에서 보이듯 중간 부분만 보여준다. 그 이유는, 그 중간부분이 실제 성장하는 메인파트이기 때문이다.
17. späte Reproduktive Phase
- GA 에서도 앞의 다른 Entwicklungsstadien처럼 내부에 성장ㅇ의 Gradient가 있다.
- Tipping (TP) : 이 단계에서 스파이크의 끄트머리가 구체화된다. 또렷. 그래서 티핑.
- Blütchenprimordia Arest & Abort : 위에 있는 녀석들은 완전히 성숙치못하고 WA, GA 처럼 멈춰서 결국 komplett absterben한다.
* 왜 다 성숙도 못할 꽃들을 더 만들까? 아마, 일단 밀은 하이브리드 종이고 이렇게 꽃을 더 많이 만들고 그 과정에 몇개는 버려라 이런 프로그램은 WT가 예전에 확보한 매우 konservative Gene의 영향이 아닐까 싶다.
* TS의 지연은 아무래도 타게스렝에, 포토페리오데에 달려있는 것 같다. 굉장히 Langtag하거나 이렇게 인공적 조건을 조성해주면 좀 더 긴 Ähre를 유도할 수 있다. 아직 분자적 증거는 없고 또한 Ts를 완전히 상실한 자연적 뮤턴트는 아무래도 만들어지기 힘든 것 같다고
* 이 사실이 시사하는 바는 큰데, 에피게네틱에서 발견한 floral mechanismen의 적용에도 불구하고 결국 매달리는 Ähre는 초기 TS에 달려있다는 것.
20. Ährenwachstumsphasen : YA에서 이미 길이는 최대에 도달, 그리고는 옆으로 막 퍼진다.
21. Ärchen und Blütenfertilität
22. Weizenertragspotenzial und Blütchen-Abort
23. Kornansatzmuster : 오른쪽 4번은 아무래도 moderne Sorte.
24. Blütchen-Abort limitiert Ertrag in Weizen : 하모요!
- fertile한 꽃만 남기거나 아니거나 (경쟁이 있다는 의미) 결국 Kornzahl에는 영향이 없었다 이말이야
25. Setentriebreduktion verzögert... :
- 여기서 봐야될 건 GA - YA에서 또는 YA - TP 에서 꽃의 숫자 자체가 확 줄어드는 Abort로 넘어가는 구간.
- 만약 Huapttrieb만 남기고 다른거 다 조지면, Blütchenabort가 확연히 지연된다.
- 그럼에도... Kornzahl에는 전후 변화가 없다... 힝
26. Blütchen-Abort limitiert Ertrag in Weizen : 이게 핵심 메시지
- 콘짤은 상관없지만, 분명히 Biomass는 detillered가 훨씬 높다는 것.
28. manuelle Seitentriebreduktion erhöht die Anzahl fertiler Blütchen am Haupttrieb durch verringerte Konkurrenz
29. erhöhte Blütchen-Fertilität :
30. Genotyp 중 퍼틸리텟 높고 그런 애들 관찰 : 7개까지 맺히는 녀석도 있다고
31. QTL auf Chr 2A : GNI2, 1
35. mRNA in situ con GNI1 in Einkornweizen : GNI1 유전자가 Blütchen pramodium 구간에서 발현됨이 발견된다ㅏ.
36. GNI1 가 꺼지면 WT에서처럼 4개의 꽃 대신 6개의 꽃이 달린다구.
38. GNI1 알렐을 통해 콘 에어트락 확 늘렸다고. 10-30%
- ohne Auswirkung auf TKG!
- 동시에 Biomasse 증가에도 영향을 줫다.
- spike number는 그럼에도 거진 차이가 없었다구.
39. 또한 거지같은 토양, 지역에서도 높은 Ertrag을 유지했다고.
40. 대부분의 T.durum 종들은 이미 95프로 GNI 유전자를 갖고 있었다고
- GNI1 은 아마 Domestikationsgen
41. 정리 : GNI1 ist ein Weizenhomolog von Vrs1 in Gerste
- Vrs1 : WT는 양쪽 꽃을 absterben,. 뮤턴트는 살려서... 이거 Gerste
42. Blütchen-Abort in Weizen : 15 Ähren * 10 Blütchen - 완전 이상적인 상황. 맥시멈!!
- 이 경우, tkg 45g, 40 t/ha!! 독일에서는 엄청 잘 자라는 곳에선 10-11t/ha
- 좀 현실적인 상황: 15 Ähren * 2.5 Blütchen -> 37 Blütchen/Ähre. 이러면 대충 10t/ha 이거 조차도 위의 독일 상황처럼 엄청 이상적인 상황
- 만약 쪼금만이라도 더 개선되서 3.0blütchen만 되어도! 12t/ha!
43. Ärchenanzahl - Weizen
44. Beeinflussung der Ährchenanzahl durch Blühzeitpunkt oder Mutationen
- 추가 Ährchen은 위에 안달리고 여기 표시되듯 아래에 달린다
* 아마, 성장이 gradient로 이뤄지기 때문이리라. 요지는, 그래디언트로 이뤄지는건 무언가의 농도, 트랜스포트가 주요펙터임을 시사
* Veredelung을 이용할 순 없으려나? 양 끝단을 연결시키면 순환을 이뤄서 더 만들수 없으려나. 또는 중력이 없는 상황에서 입체적으로 식물을 기르기 위한 디자인이 될 수 도.
- 미라클 위트 : 몇안되는 TS 뮤턴트를 이ㅜ러낸 경우.
45. 분더바이쩬의 비교!
- 딴건 거의 다 퐈지티브했으며 TKW 는 15프로 적었지만, 뭐 이건 그렇게 Ertrag 에 주요하진 않다!
46. 유전자 찾으려고 Gerste Homolog 랑 비교해봄
- Com2
- 디플로이드랑 헥사플로이드 간 뮤턴트의 퍼틸리티 문제 발생 상황이 달랐다! 이게 시사하는 바는 Polyploidisierung이 퍼틸리텟에 특정 역할을 하지 않을 까 하는것.
- Verzweigung과 Fertilität 의 연관관계를 유전자 단위에서 관찰
47. Sequenzieren von Weizenmutanten : Wunderweizen의 Verzweigung & Fertilität을 책임지는 유전자를 찾기위함
48. Phänotypen in F7 derived RILs : 왜 7세대냐 했냐면 이러고 나니 온갖 미친 페노튭이 생겼기 때문.
- 그 이유는 몰까? 다른 유전자가 있거나, modifier 가 있거나 그리고 나는 이해가 잘 안가거나
- 그 이유가 이 뮤턴트 bh t 는 rezessiv Mutnat - A1 (크로모좀 2A) 이었는데 B1 에서도 비슷한 bh t 즉 도 다른 크로모좀 B2 에 있는 알렐도 찾았다!
* 7세대나 했던 이유 : 우선은 Homozygote만들기 위한것. 하지만 여러 표현형이 나왔다는 건 실패했으며 동시에 다른 알렐이 들어있다는 것. 하지만 이럴거면 분명이 좀 더 초기 세대에서 실패가 자명해졌을 것 같은데 왜 굳이 7세대를 끌고갔는지는 모르겟다.
- aa 가 bb보다 훨씬 강한 효과를 가진다는 것을.
49. Mendelization of the miracle wheat allele : 찾은 미라클을 륙크로이쭝을 통해 여러 세대를 거쳐 궁극적으로 NIL 더 많은 Verzweigung 가진 녀석을 얻어냈다.
- 왜 도너처럼 미친듯이 안달릴까? 다른 유전자가 있기보다 이들을 repressor하는 유전자의 존재를 시사할지도.
그다음 수업 : Mais, REis 등 다른 종에서 Meristen 과 Ärechnentwicklung in Gerste 를 다룰거임!
- 교짱은 비오팜에서 프락티쿰하다가 좋아서 농학 공부. 석사로 랍스 genetische Identifikaiton (Vererbung 테스트) 했는데 결과는 영 아니었다고. 그러다가 비오테히놀로기 히비하다가 관심생겨서 레지스텐쯔가고 공부 방향으로는 농장에서 일을 하기가 힘들고 또 내 농장이 있어야하는데 그렇진 못해서 현실적 타협을 했다고.
- 오늘의 명언 : alle Morphologie hat Grund
[아마 마지막 시간]
1. 여러 메리스템 - Reis
- Kurztag PF. Rachilla가 밀 이런거에 비해 매우 unterdrückt
- Branch und Spikelet : 핑크가 스카이클메리스템. 초록색이 브랜치(Verzweigung) 메리스템.
- APO1-D1 : reproduktive Meristem 단계에서 지연시키는 듯.
2. Mais : Auxin이 난리도 아니여. 옥신과 Blütenmeristem 의 관계가 이 뒤를 위한 기본
- 옥수수는 맨 위에 수꽃, 그 아래 암꽃이 피나보다.
- Tassel은 그 시작점의 Branch Meristem덕분에 수꽃이 여러갈래로 자라는 것임.
- Blütchenentwicklung und Geschlechtdifferenzierung : 보면 알겠지만, 옥수수는 초기에 둘 다 양쪽 성을 다 가질 가능성이 있는 상태인데 그게 시간지나면서 abort 되면서 성 분화가 한 개체내에서 발생.
- RAMOSA gene : 뮤턴트. Verzweigungsmeristem이 단순히 꽃 뿐만아니라 Korn에도 영향을 미친다.
* 이런 곧은 와일드 타입의 형성에 가설 중 하나는 농업환경과 Beschattung. 좁은 곳에 밀집 Anbau하면서 에너지를 더 효율적으로 해야된다. tassel에서 Pollen만드는 것도 어ㅁ청난 에너지. 이 에너지를 콘에 쓸 수도 있겠지. 그리고 어차피 eng하니까 폴랜이 엄청 많을 필요도 없다.
3. Gerste : 왜 한개, 3개 이렇게 나는데 zweizeilig, sechszeilig? 앞뒤로 자라니까 2배되서 각각 2줄 6줄이 된다.
- Zeiligkeitsmutanten : Entwicklungsgradient 와 관련있다! vrs2, vrs3 을 비교하면 그라디엔트가 반대로 작용. 그리고 하나 더 알건 이 모든 유전자들은 negative Regulator (WT에서는 Sterilität 을 일으키는거지)
* 2 줄 보리는 Brauprozess 에 유리? 6줄에 비해 더 호모게니텟하다?
- Korngrösse : 초록색이 메인콘, 파란색이 레터럴. 빨간색은 더 달린 녀석 (6줄 이상)
- Zeiligkeit 의 보리에서 중요성 : Kornzahl (2,6), 그리고 컴비네이션 뮤턴트가 보여주듯 Korngroß
- spike development vrs4 : 사진을 보면 determiniert 되지않고 계속 연장되어 자라남을 알 수 있다.
- ZmRAMOSA2 가 Vrs4 : 아따 결국 찾았네요
- 그러고 나면 이제 구체적으로 조직 어디에서 이 유전자가 발생하나 :
- Vrs4 가 Vrs1 를 regulieren 한다, upstream으로.
4. Grain number : 제일 아래는 각 단계마다 발달하는 agronomische Eigenschaft
5. Gerste - Ähfferenzierungsphase : 2번째 단계에서 Entwicklungsgradient발생.
- Mikrostadien 을 커비 스케일 책에서 확인할 수 있다 : 우리 프락티쿰에서 할 것.
* 밀의 특징은 저 3 mound 가 발견된다는
- Waddingtion scale : 이것도 Mikrostadien 스케일. 1-10까지 있으며 보다시피 매우 fein하게 분류가능. keine Hexerei!
- 커비와의 차이를 보자면, white anthere stage 를 웨딩턴은 이렇게 구분하지만 커비는 그냥 한 스테이지로 퉁친다.
* 시험에서 각 구조 기억하는게 중요.
6. VRS2 : 마지막 보리 뮤턴트. GA 와 연관. 아마 목적이 중간 부분이 다른 부분보다 Ähre가 빨리 꽃이 달리도록 하는 목적으로 활처럼 농도가 퍼지는 듯.
- positiver Regulator der Auxinbiosynthese
[Wurzelentwicklung und -achitektur]
1. Rhizosphäre : unmittelbar durch eine lebende Wurzel beeinflussten Raum im Boden
- 매일 사람이 소비하는 영양은 3.5톤 정도의 흙에 있는 미네랄을 소비
- Makro / mikro 미네랄의 기준 : 보통 Mangel이 발생하지 않는 최저 Schwelle를 의미
2. 뿌리는 verborgene Hälfte der Pflanzen 이다. 거의 똑같이 반을 차지하는 경우가 많은듯
3. 모든 Gräser = monokotyl ( Hauptwurzel 이 메인으로 발달하고 Lateral 은 좀 적게, 이 둘이 메인딜러)
- dikotyl : Hypokotyl 에서 Crown Root, Brace root 가 영양 흡수 메인 딜러. Allorhizo - 겉으로 보면 모든 뿌리가 CR 로 같은 것처럼 보인다고.
* Phytomer : 모든 Phytomer는 아래 RAM 갖고 있는데 이게 Veredelung이 가능한 이유 중 하나. 여기서 CR 이 쇽쇽 나올 수 있다고.
- At : Wurzelhaube 가 겔에서 기르면 안나올 수도.
4. Wurzelhaar : 매우 강하게 성장한 하나의 세포다!! 즉, vaskulares System 없고 단지 영양 흡수, 전달 등에 관여한다고.
5. Zell typ-spezifische At Marker Linien : 뿌리에서 구역마다 특정 Promoter가 작동하는 구역이 나눠져있다고. 즉, 이 마커로 뿌리의 지역을 구분할 수 있다
- Konforkal Mikroskopie (단백질 염색해서 관찰) 또는 Durchflusszytometire (FACS,
6. 식물 seitenwurzel 이 자라려면 Epidermis 세포벽을 무너뜨려야 되는데 세포벽 녹이기 위한 효소를 만들어내면서 왜 본인의 세포벽은 안녹을까?
- junge Wurzel 은 메틸화되어있다! 그래서 Säure 에 안녹는다고.
- 세포벽은 Zucker 로 카르복실, 에틸화되어 있다구. Ca2+ 가 들어오면 라둥가진 벽 분자들을 더 강하고 잡아서 딴딴해진다고.
7. Funktion der Wurzel : Halt, Wasseraufnehme, Nährstoffaufnahme + Speicher, Kohlenstoff-Pumpen (이걸 내서 주변 미생물을 모집, 주변 영양을 흡수가능하게), Wachstumsregulation und Entgiftung + Sensing (땅의 온도는 상대적으로 Spross보다 안정적이기에 이걸 기반으로 감지)
8. 뿌리성장을 결정하는 요소 2가지 : Genotyp + Standort
- 온도와 어린 뿌리 : 어린 시절에는 땅의 온도가 뿌리 성장에 제대로 영향미침. 즉, 빛 외에 주변 식물과 맞춰줘야 하는 펙터로써 온도를 변수로 고려해야 한다는 것.
- 식물을 땅에 심을 때 다져주는 거 : 눌렀나 안눌렀나가 향후 식물 자라는 데 장기적 영향
- 질소부족과 뿌리성장 :
- 뿌리 성장은 lokal System : 지엽적으로 Reaktion
9. 여기의 목표 : 여러 환경에 optimal하게 반응하는 Wurzelsystem 을 찾거나 만드는 것 (Züchtung)s
-
- Phänotypisierung : 살아있는거 옆에 그대로 파서 관찰. Feldbediungung. sehr aufwendig. 그래서 hydroponisches System으로 물에다 길러서 관찰. 물론 이건 자연적인 환경과는 거리가 멀다구. 또 다른건 Löschpapier-system 이라고 종이에다 기르는거. 제일 쩌는건 Computertomograpie 라고.
* 브롬 Bor 는 세포벽 구성에 핵심 요소인데 이게 부족하면 뿌리 Differenzierung 이 잘 안된다. 그리고 Bor 는 ungeladene 라서 물에 녹아야 흡수 가능한데 Bor가 있어도 trockenheit 때문에 Bormangel이 발생할 수 있다. 예를 들어 Raps 처럼!
- 매우 열정적이고 실험 페노타이핑 등에서 효율을 추구한다. 특히 공학적 적용을 고민한다면 Gene 보다 Function에 집중해야 한다는 메시지가 매우 인상적이었다,
[ molecular bases of Apomixis in Hypericum perforatum - Johanniskraut]
1. 6 Faktor : active compounds (보통 이게 apomitic한지 모른다고), diversity, Development (Apomixis), Transcriptomics, Genomics, Metabolomics
2. 식량과 관련된 현상황 : Yield 증가는 꾸준히 발생했지만 현재 한계점에 도달한 듯 보인다.
3. Apomixis : asexual reproduction through seed 장점이 뚜렷한 반면, Not in major crops!
- Progenies are clones of the mother, instant fixation of superior phenotypes (heterosis)
- 특정 교수들 말론 쌀에 적용하면 처음 5년만에 800밀리언 예상한다고
4. Apotechnology : unreduced gametes, parthenogenetic Ebryo formation, active Endosperm
5. Gemetophyte, Sporophyte,
6. sexual female Gametophyte Development :
- MegasporoGenesis : archesporial cell -> Megaspores -> functional Megaspore
- Megagametogenesis : Antipodal (3n, 아직 역할을 잘 모른다고!), Central Cell (2n), Egg cell (n)
- polygonum : ?
7. What about APOMIXIS : 종류는 sporophytic APO (Citrus 에서 발견, 완전 골때림. 그냥 체세포가 씨앗이된다고?), gametophytic APO (Hypericum)
- 과정은 앞에말한 3가지 조건.
- Drews and Koltunow, 2011 : 이들의 논문이 아마 무성생식 식물을 가장 잘 설명한 것 같다고
- Diplospory 의 특징 중 하나 : Megasporogenesis (피메일 가메토) 반들어지는 중 옥타플로이드 되고 개판인데 보통은 이러면 레탈인데 씨트러스는 이걸 허용해서 자란다고
- aposporous initial : Hieracium 이 그 예시. mitosis 로 만들어지고 mitosis 로 각 1개의 가멧토가 만들어지는데 미토시스로 만들어진 건 사라진다고.
8. Hypericum perforatum : 여러 의학적 효과
- Genotype에 따라 3가지 가능성 : 섹슈얼, 아포믹스, 둘 다 (마치 렌덤하게 Ratio하는 듯)
- 섹슈얼은 디플로이드 또는 헥사플로이드, 아포믹스는 테트라
9. Addressing apomixix : 크게 5가지 방향
- Happy locus, Development (morphology) : 이 두가지가 주요하게 역할한다고.
- Development (transcriptomics), Genomics (genome sequencing), Genomics (GBS)
10. Hypotheses of Apomixis Hypericum : sexucal ancestors -> Hybridization -> Genome collision -> expres. tip regulators disrupted -> dev. functions disrupted -> Apomixis (emergency procedure 아마!)
- 앞서 설명한 hyragium 에서 아포포러스 설명한 생식세포와 아포믹스 세포 경쟁이 존재하지 않고 단지 아포믹스만 발생.
11. 최종단계까지 가더라고 씨앗이 fertility를 가질 가능성은 섹슈얼보다 낮다. 즉 Abortion 이 높다 !
12. 이녀석의 aposporows 에는 meiosis 1 만 있고 meisis2 는 없다. 그 다음에는 Mitosis,
13. 파괴없이 아포믹스 스크리닝 : Flow Cytometric Seed Screen (FCSS) : 이 방법을 볼땐 ploidy ration 를 확인하는게 핵심. Embryo:endosperm 이 1:1.5 이면 보통 섹슈얼,
* Polycomb, PRC2 이 FCSS 에 주요한 유전자임.
14. probable epigenetic : IBM1 (H3k9 demetylation)
15. Genotyping by Sequence : 전부 씨퀀싱해서 지노타입 잡는건 너무 비싸니까 숏컷.
* 아포믹스 구루 : Elvira Hörandl, Koltunow, Grossniklaus, Sharbel (이페카에 있음)
'독일 석사' 카테고리의 다른 글
| [molekulare Marker in der PFzüchtung] (0) | 2019.10.30 |
|---|---|
| [Selektion in der PFzüchtung] (0) | 2019.10.18 |
| Schreiben - Seminar, Protokoll, Masterarbeit (0) | 2019.09.17 |
| [Epigenetik] (0) | 2019.04.26 |
| [Ökologischer Landbau] (0) | 2019.04.16 |